INTRODUCCIÓN
La competencia entre plantas es un proceso ecológico importante, tanto en la comunidad de un tapiz natural, como el que se establece en una población de plantas cultivadas. La competencia en la comunidad, puede ser considerada tanto a nivel de parte aérea como a nivel radicular (Grace y Tilman, 1990; Wilson, 1988).
En 1869 Ernest Haeckel utiliza por primera vez el término ‘ecología’ como referencia al estudio de la interrelación entre organismos y el ambiente (Begon y col., 2006). El ecosistema como la unidad funcional de estudio, donde interaccionan factores bióticos y abióticos, fue la definición propuesta por primera vez por Tansley (1935). Relacionados estrechamente a la noción de ecosistema se define el hábitat y el nicho ecológico. El hábitat es el lugar físico del ecosistema, donde una especie se encuentra adaptada, en tanto que el nicho ecológico lo constituye la relación del individuo con los factores bióticos y abióticos de su ambiente (Rejmanek y Jenik , 1975).
Se define como una población al conjunto de individuos de la misma especie que comparten el mismo hábitat, en donde la dinámica de los cambios está determinada por los procesos de nacimientos y muertes. El estudio de la abundancia de la población y cómo evoluciona en el tiempo es el aérea de investigación de la ecología. (Colyvan, 2008; Berryman, 2002) La permanencia de una especie en un ambiente dado indica su capacidad de competir con otras especies de hábito y requerimientos similares por los mismos recursos abióticos. Además, los aspectos anatómicos, fisiológicos y bioquímicos permiten por una parte, reducir la competencia directa y por otra, aumentar la eficiencia en la utilización de los recursos luz, agua y nutrientes (Medina, 1996). La población puede fluctuar en el tiempo debido a la disminución de recursos o puede sufrir variaciones debido a la presión biótica de otra especie que está incrementando su número de individuos. Las poblaciones están limitadas por los recursos en su capacidad de crecer y para una especie dada, el número de individuos que un determinado hábitat puede mantener, se define como capacidad de carga (Colyvan, 2008). De acuerdo a Bazzaz (1996) y Odum (1953) la interacción Planta-Planta puede ser positiva, neutra ó negativa. Luego Odum (1953) define cada una de estas situaciones de interacción como:
Neutras: cuando no existe ningún tipo de interacción entre las diferentes especies.
Positivas: cuando se favorece o se beneficia alguna especie.
Negativas: cuando se perjudica alguna especie (competencia)
La vegetación natural, compuesta por dos ó mas poblaciones en un mismo nicho ecológico, mantendrán una competencia ínter específica por los recursos del ambiente, lo cual afectará en diferentes grados el crecimiento y/ó supervivencia de cualquiera de los componentes del tapiz vegetal (Matteucci y Colma, 1982).
La eficiencia en el uso de nutrientes es un aspecto clave en la evolución y en el éxito competitivo de las especies. La competencia ínter e intra específica en el ambiente por el recurso fósforo, puede generar una serie de respuestas de plasticidad de adaptación de las plantas al hábitat considerado. Las especies que evolucionan en ambientes restrictivos en determinado recurso tienden a desarrollar mecanismos que aseguren la competencia por el mismo. Si bien la competencia subterránea ha sido menos considerada en el pasado próximo debido a las dificultades para su estudio, los avances en los últimos años han demostrado que la competencia de las raíces por recursos es más intensa e involucra una mayor complejidad que la competencia aérea por la luz (Casper y Jackson, 1997). El suministro de P en la parte aérea de la planta inhibe el desarrollo de mecanismos a nivel de raíz que están implicados en el incremento de la absorción de fósforo desde el suelo. Esto indica una interdependencia parte aérea-raíz que involucra señales de suficiencia interna de fósforo en la planta (Marschner y col., 1986).
COMPETENCIA ENTRE PLANTAS
La competencia es definida como la interacción negativa entre individuos por un recurso determinado y limitado, que tiene como consecuencias un efecto sobre el crecimiento y/o reproducción de los mismos en su hábitat (Bullock, 1996).
La competencia entre especies será mayor cuanto más semejante sean entre si sus requerimientos por los recursos del hábitat. En tanto que la población sobrepasa la capacidad de carga de ese hábitat, los recursos se agotan más rápido de lo que pueden ser renovados, por lo que unas poblaciones pueden prevalecer sobre otras. Es así que la competencia radicular, en un volumen dado de suelo afecta el crecimiento, disminuye la supervivencia o la fecundidad de las plantas vecinas debido a la reducción de los recursos disponibles del suelo.
Golberg (1990) considera la competencia, desde el punto de vista de la interrelación de las plantas por un determinado recurso, a la relación entre la manera que este recurso es afectado y la respuesta de las plantas a las variaciones del mismo (Fig. 1). Para que se establezca una competencia, a nivel radicular, un individuo debe ejercer un efecto negativo sobre la disponibilidad de uno ó más recursos, a los que otra planta presenta una respuesta positiva, como por ejemplo el crecimiento, la supervivencia o la reproducción (Goldberg, 1990; Grace y Tilman, 1990).
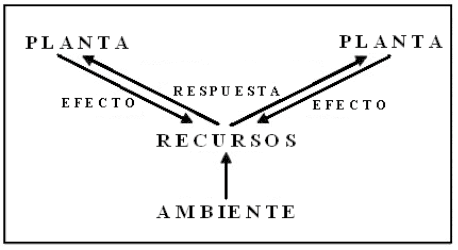
Figura 1. Esquema en el cual una planta ejerce un cambio sobre un recurso del ambiente y la otra planta en competencia, responde al cambio. Tanto el efecto como la respuesta deben contribuir a establecer la competencia. (Casper y Jackson, 1997)
En un mismo hábitat, las relaciones Planta-Planta pueden ser neutras para un recurso dado y negativa para otros recursos en el mismo momento. La capacidad competitiva no está necesariamente correlacionada. Por ejemplo, una planta puede mejorar la absorción de agua a través de un sistema radicular más profundo, aprovechando así una fuente de agua no disponible para las plantas vecinas de raíces poco profundas. En este hábitat no se incrementa la competencia por el recurso agua, incluso podría considerarse que disminuye, pero si puede incrementarse la competencia por nutrientes minerales o por la luz, como consecuencia de un crecimiento más vigoroso de las plantas o aumento de la densidad de plantas. Del mismo modo, la sequía puede aumentar la mortalidad denso-independiente de las plántulas, simplemente porque sus raíces no están bien desarrolladas. Las plantas también difieren en su capacidad de metabolizar e integrar los recursos del suelo a la biomasa, lo cual introduce el concepto de eficiencia del uso del agua o la eficiencia del uso de nutrientes (Nobel, 2009).
La habilidad competitiva de las especies depende del ambiente y el momento considerado. El nivel de nutrientes en el suelo determina la capacidad de competencia y como consecuencia la composición botánica. En una pradera mezcla de gramínea (Festuca arundinacea cv. Demeter, Lolium perenne cv. Kangaroo Valley y Phalaris tuberosa cv. Australian Commercial) y trébol blanco (Trifolium repens cv. Grasslands Huia), en suelos con deficiencia de fósforo, la capacidad de competencia de las malezas nativas (Hypochoeris radicata y Eragrostis trachycarpa), determinó que las mismas prevalecieran en el cultivo, principalmente en el cuarto año del experimento (Fig. 2). Con altos niveles de fertilización fosfatada la gramínea domina sobre el resto de los componentes de la pradera, lo cual se hace notorio en el cuarto año de establecido el cultivo (Wolfe y Lazenby, 1973).
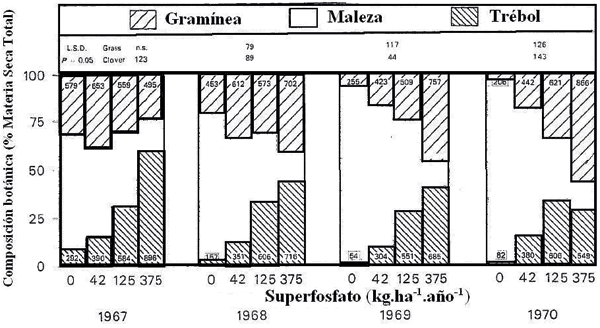
Figura 2. Respuesta a la aplicación de superfosfato, durante cuatro años en una pradera de gramínea y trébol blanco (Wolfe y Lazenby, 1973).
CARACTERÍSTICAS RADICULARES DE LA CAPACIDAD DE COMPETENCIA
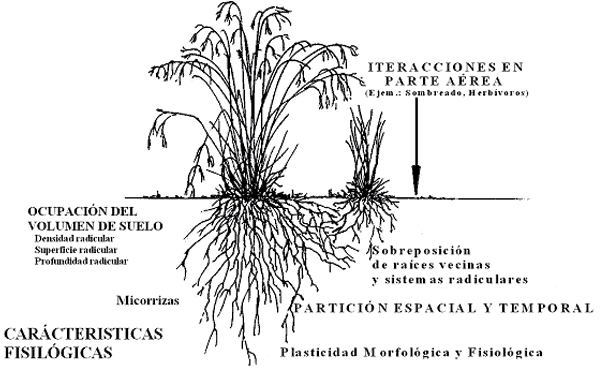
Figura 3. Características morfofisiológicas de las plantas que contribuyen en la capacidad competitiva a nivel radicular (Casper y Jackson, 1997)
Es necesario comprender los mecanismos morfológicos, fisiológicos y bioquímicos involucrados en la captación de los recursos del suelo, mecanismos que promueven la competencia por esos recursos, favoreciendo la capacidad competitiva de la rizósfera (Fig. 3).
La capacidad de exploración del suelo por las raíces y por tanto su capacidad de competir y/ó coexistir de las especies depende de la velocidad de crecimiento, la densidad de raíces, el diámetro, la superficie total de absorción y la profundidad de exploración radicular. A su vez la complejidad del sistema, hará que toda variación en la parte aérea tendrá consecuencias en la actividad radicular, por ejemplo variaciones en la fotosíntesis ó en la translocación de fotoasimilados.
Trabajos realizados por Caldwell y col. (1991) muestran que la densidad de las raíces de tres especies (Artemisia tridentata, Pseudoroegneria spicata y Agropyron desertorum) en competencia, no explica por si misma las diferencias en la absorción de nutrientes, en tanto que ha sido demostrada la importancia de los pelos radiculares en las diferencias de absorción de P por las plantas (Föhse y col., 1991; Itoh y Barber 1983ª; Caradus, 1981) debido a la capacidad de incrementar la superficie de absorción de nutrientes (Claassen, 1990). Trabajos realizados con seis comunidades de plantas, cuatro como monocultivos y dos como mezclas ( Festuca ovina L. y Achillea millefolium L. o
Phleum pretense L. y Trifolium pratense L.), demuestran que existen diferencias en profundidad de exploración de las raíces y la capacidad de absorción de nutrientes, pero ese comportamiento depende según se encuentren como monocultivo ó como mezcla (Jumpponen y col., 2002). Steingrobe y col. (2001) en trabajos realizados con plantas de cebada, remarcan la importancia de considerar la dinámica entre la muerte y crecimiento de nuevas raíces. La generación de nuevas raíces es beneficiosa en cuanto incrementa la superficie activa de absorción, pero este hecho dependerá del balance raíces nuevas y viejas y la capacidad de exploración.
Al estudiar las competencias de las raíces por los recursos del suelo, debe considerase que los elementos minerales entran en contacto con la superficie de la raíz por tres procesos: 1. intercepción de los nutrientes, 2. flujo de masa y 3. difusión en el suelo (Marachner, 1986). En la naturaleza el flujo de masa puede ser considerado como el proceso más importante de suministro de nutrientes a la raíces, pero es relativamente poco importante en el caso de los nutrientes de baja movilidad ó cuando la disponibilidad es limitada (Chapin, 1991). La absorción de iones por intercepción de la raíz puede representar menos del 10% del total y puede ser considerado el proceso de menor importancia de los tres mencionados (Marschner, 1986). El proceso de absorción de iones minerales depende del elemento considerado, así el nitrógeno (N) llega a las raíces a través de la combinación de procesos de flujo de masa y de difusión en tanto que la absorción del fósforo inorgánico (Pi) depende en gran medida de la intercepción por la raíz (De Willigen, 1986). El coeficiente de difusión del NO−3 es de 1 × 10−10 m2 s−1 (Barber, 1984), en tanto que para el fósforo el coeficiente de difusión en agua es de 0.89 x 10-5 cm2 s-1 y en el suelo se tienen valores experimentales que dependen del tipo de suelo y cuyos valores oscilan entre 1×10- 6 a 4 × 10−15 cm2 s−1 (Schenk y Barber, 1979; Barber, 1984; Barber, 1986). El proceso de difusión suele ser el paso limitante en la absorción de los nutrientes por las raíces de las plantas (Fig. 4) (Nye, 1966; Chapin, 1986). En consecuencia el P está menos disponible en el suelo para absorción por las raíces que el N y en una situación de competencia de las raíces, el ión menos móvil en el suelo será el que se agote más rápido y establezca la condición de factor limitante y por tanto la competencia por el mismo.
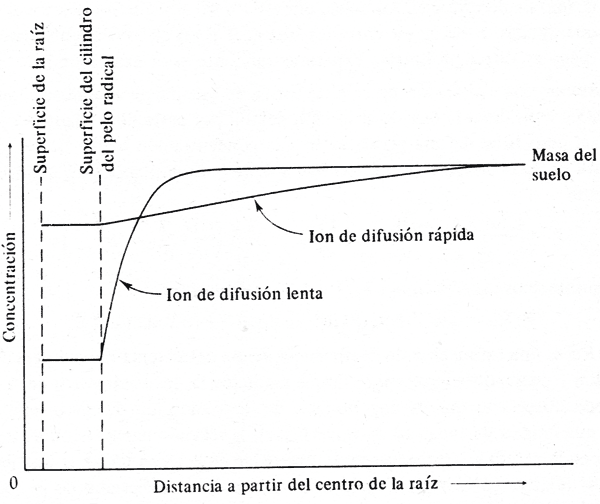
Figura 4. Efecto de la velocidad de difusión del ión sobre la disponibilidad del mismo en la superficie de la raíz (Nye, 1966)
FOSFORO
La deficiencia de fósforo inorgánico (Pi) es uno de los principales factores que limitan el crecimiento y la productividad vegetal en muchos de los ecosistemas. La mayoría de los suelos poseen baja disponibilidad de fósforo lo cual puede inducir a las plantas que compiten a desarrollar mecanismos que facilite la absorción del nutriente.
La supervivencia de las plantas y el rendimiento de los cultivares, en condiciones de deficiencias de Pi se han relacionado muchas veces con la arquitectura de la raíz (Vance y col., 2003; Raghothama, 1999). Son muchos los factores asociados a la raíz que facilitan la competencia por el Pi los cuales incluyen la plasticidad de adaptación y morfología de la raíz, superficie de absorción, arquitectura, densidad y longitud de pelos absorbentes, velocidad de absorción, capacidad de modificación de la rizósfera y asociaciones micorríticas. La competencia subterránea a menudo disminuye con el aumento de los niveles de nutrientes (Casper y Jackson 1997; Raghothama, 1999). Es en estas situaciones de alta disponibilidad de fósforo en el suelo, que pueden no expresarse los mecanismos de competencia
CINÉTICA DE LA ABSORCIÓN DE FOSFORO POR LAS RAÍCES
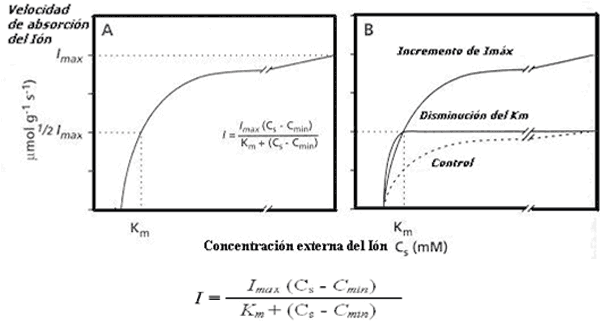
Figura 5. A.- Relación entre la absorción neta de Pi (I) y la concentración en la solución externa del ión (Cs)
B.- Cinética de la absorción en plantas control y plantas creciendo en escasez de Pi en las cuales se inducen diferentes sistemas de regulación de la absorción
En el caso del ión Pi, si el proceso de difusión no es limitante, la absorción del mismo por la raíz se incrementa en respuesta al aumento de concentración en la solución del suelo, hasta alcanzar una velocidad máxima constante ó de saturación, la cual es independiente de la concentración de iones de la solución externa (Fig. 5A). Esta relación de flujo de ingreso de iones a la raíz, en función de la concentración en la solución externa, es descrita por la función de cinética enzima-sustrato de Michaelis-Menten modificada (Epstein, 1966; Nielsen y Barber, 1978).
La absorción neta de iones (I) es función de la velocidad máxima de absorción (Imáx) a la mayor concentración del ión en la solución externa (Cs), la concentración mínima en la cual el ingreso neto de iones a la raíz es igual a cero (Cmin) y la afinidad del sistema de transportadores por el ión (Km) cuyo valor es 0.5 Imáx . Los parámetros de la función de absorción de nutrientes Michaelis-Menten, determinan las diferencias genéticas entre poblaciones o de adaptación a condiciones de escasez del ión en la solución. Así Imáx es un parámetro que tiene su utilidad en describir la capacidad de la raíz en la absorción de iones, la constante de Michaelis-Menten (Km), representa la capacidad de una raíz de absorber a bajas concentraciones de sustrato ó la afinidad del sistema transportador por el ión. El parámetro Cmin representa el balance entre el ingreso del ión a la raíz mediado por transportadores y el egreso de ese ion contra un gradiente de potencial electroquímico, por debajo de esa concentración de nutrientes, no existe absorción neta de iones hacia el interior de la raíz. Considerando diferentes especies ó cultivares (Fig. 5B) puede observarse que para sistemas de diferente afinidad se produce una adaptación al disminuir la concentración externa del nutriente, a través de un cambio en el Km ó un incremento de Imáx lo cual tiene como consecuencias incrementar la capacidad de absorción de fósforo de la solución del suelo (Marschner, 1986).
INTERACCIONES ENTRE RAÍCES.
Además de responder al gradiente de recursos generados por las raíces vecinas, también se dan respuestas a efectos directos producidos por otras raíces. Los mecanismos directos de interacción entre raíces incluye el exudado de toxinas que difunden en la rizósfera, exudados químicos no tóxicos y señales eléctricas (Schenk y col., 1999).
Cuando se considera la competencia de las raíces por el nutriente fósforo, puede observarse que los valores de velocidad de absorción estimados son superiores a los valores determinados experimentalmente. Se establece una relación curvilínea entre la absorción de fósforo y la densidad de raíces, debido al solapamiento de zonas empobrecidas en el nutriente fósforo. Esta competencia debe ser considerada cuando se intenta relacionar la densidad de raíces en los diferentes horizontes del suelo y su contribución al suministro de fósforo (Fig. 6 y 7) (Nye y Tinker, 1977; Itoh y Baber, 1983b)
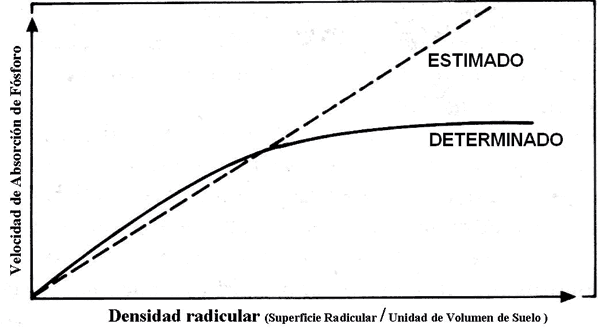
Figura 6. Relación entre la densidad radicular y la velocidad absorción dependiendo de la difusión del nutriente fósforo
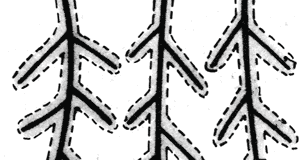
Figura 7. Zonas de agotamiento de nutrientes, cuando la velocidad de difusión del elemento considerado es baja
Un mecanismo indirecto se establece en el sistema radicular, en competencia por los nutrientes del suelo, entre las raíces del sistema radicular de la propia planta o de sistemas radiculares de plantas vecinas (Fig. 8). La velocidad de absorción estará entonces determinada por la distribución de las raíces y la velocidad de agotamiento del nutriente en el suelo (Nye, 1979).
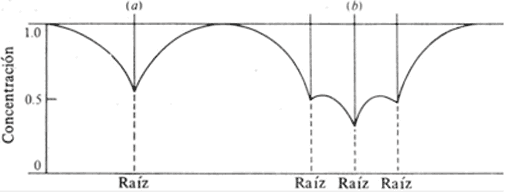
Figura 8. Concentración de iones alrededor de (a) una raíz aislada y (b) tres raíces vecinas, donde se observan las zonas de agotamiento (Nye 1979)
Los trabajos precedentes, determinan la densidad de plantas para un hábitat determinado, que dependen de parámetros como la longitud de la raíz, superficie de la absorción por unidad de volumen de suelo y el patrón espacial entre raíces.
Baldwin y Tinker (1972) trabajando con especies en competencia, (Allium cepa, Agropyvon repens, Polygonum aviculare, Brassica napus y Lolium multiflorum) (Fig. 9) demuestran que cuando se establece una competencia interespecífica se modifican los patrones de crecimiento de las raíces tanto en profundidad como en la arquitectura de la mismas.
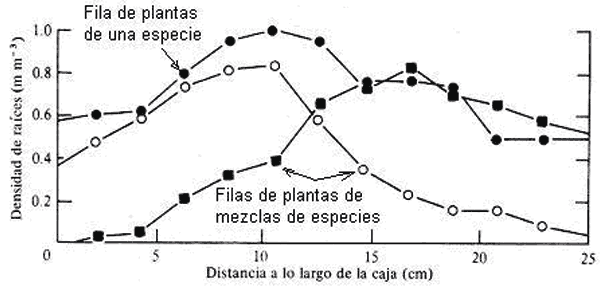
Figura 9. Cambios en la densidad de raíces al variar la distancia a lo largo de la caja de plantas en filas de una especie y filas de diferentes especies creciendo conjuntamente (Baldwin y Tinker, 1972)
Determinado los parámetros de cinética de la absorción de Pi con plantas de soja en varios tipos de suelos, los resultados concuerdan con los obtenidos utilizando modelos de simulación (Fig. 10). En situación de alta concentración de P, la absorción se realiza en mayor proporción por la velocidad de elongación de las raíces (k) y por el diámetro de las mismas (r0) y tienen una importancia menor los cambios en los parámetros de la cinética de absorción (Km, Imáx y Cmín). Lo inverso ocurre a bajas concentraciones de fósforo en la solución del suelo. Los factores del suelo son más importantes cuando el nivel de fósforo aumenta que cuando disminuye (Coeficiente de difusión De y poder buffer del suelo b). En cualquier valor de rango de variación estudiado, la transpiración (v0) no ejerció ningún efecto en la velocidad de absorción. En condiciones de baja densidad de plantación, no se observa competencia entre raíces (ri). Para este caso de plantas de soja puede concluirse que para un nutriente relativamente inmóvil como es el fósforo los parámetros cinéticos son importantes a bajas concentraciones en tanto que los morfológicos lo son a altas concentraciones.
La difusión no resulta ser un factor limitante cuando la demanda de la planta es baja ó cuando la concentración de Pi en la solución del suelo es alta (Drew y Nye, 1969)
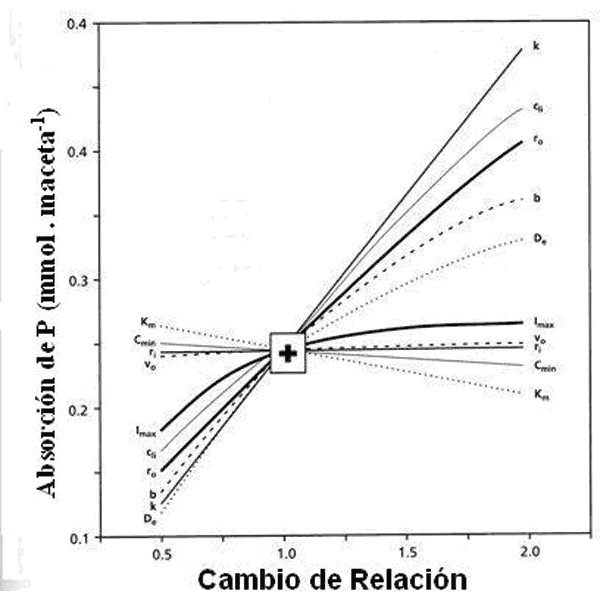
Figura 10. Cambios en los parámetros de absorción de P según modelo matemático, para plantas de soja creciendo a diferentes concentraciones de P (Cji) . (Silberbush y Barber 1983)
MORFOLOGÍA DE LAS RAÍCES, COMPORTAMIENTO EN LA COMUNIDAD
La competencia de las raíces por los recursos en el perfil del suelo, está determinada por las diferencias genéticas de las especies y variedades, los aspectos biológicos del suelo, las propiedades físicas y químicas y las variaciones estacionales del medio ambiente (Pearson, 1965; Reuss y Danielson, 1975; Taylor , 1983; Klepper, 1987).
Humphries y Bailey (1961) estudiaron los hábitos radiculares de ocho especies de tréboles anuales, principalmente trébol subterráneo (Fig. 11). El suelo utilizado fue arenoso, sin límite de profundidad y sin ningún cambio abrupto de textura u horizonte compactado que impidiera el libre desarrollo de las raíces. Observaron que las especies poseían diferentes hábitos de desarrollo radicular. El momento de floración estaba asociado a la profundidad de exploración de las raíces. Las variedades de floración más tardía poseían sistemas radiculares más profundos que las variedades de floración temprana. Como consecuencias de estas diferencias, las variedades de floración más tardía produjeron más forraje y presentaron una mayor resistencia a condiciones de stress hídrico en los meses de verano.

Figura 11. Distribución de raíces de ocho tréboles anuales en un perfil homogéneo de suelo (Humphries y Bailey, 1961).
No obstante las diferencias genéticas de desarrollo radicular, las diferentes especies ó variedades pueden presentar plasticidad morfológica y fisiológica. Esta capacidad de realizar ajustes ante los cambios locales del ambiente suelo, puede ser crítico para el éxito de la competencia subterránea de la planta considerada.
Ventajas de la plasticidad pueden ser consideradas en los términos de cuanto más nutriente como P adicional es absorbido por las raíces y cuan rápido, pues el aumento en la velocidad de absorción puede ser muy importante ante la presencia de raíces que compiten por el mismo recurso. Muchas plantas responden espacial y temporalmente a los cambios en la concentración de fósforo en el suelo (Fig. 12). Cambios en la proliferación de raíces en la zona de mayor concentración del nutriente permite a la planta aprovechar las condiciones de heterogeneidad de Pi en los suelos (Drew y Saker , 1978).
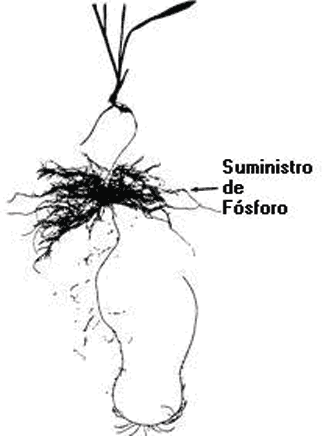
Figura 12. Distribución de raíces de una planta de cebada cultivada, parcialmente expuesta a una elevada concentración de Fósforo, mientras que el resto se expone a baja concentración (Drew y Saker, 1978).
Fransen y col. (1998), estudiaron la distribución del P en el suelo, su impacto en la biomasa radicular y la absorción de nutrientes, por las gramíneas Lolium perenne L., Holcus lanatus L., Festuca rubra L., Anthoxanthum odoratum L. y Nardus stricta L. Las especies consideradas produjeron más biomasa radicular en los suelos donde el nutriente se distribuyó en forma heterogénea, que en aquellos suelos en los que el nutriente se distribuyó en forma homogénea. Así mismo, en los tratamientos de distribución heterogénea del P, se encontraron diferencias significativas entre las especies para la longitud radicular y la densidad del sistema radicular, registrándose los mayores valores para las especies de rápido crecimiento como L. perenne, H. lanatus y F. rubra. Estos cambios en el sistema radicular no se correlacionaron con la absorción del nutriente fósforo, por lo que un mayor desarrollo radicular no implica necesariamente una mayor absorción del nutriente.
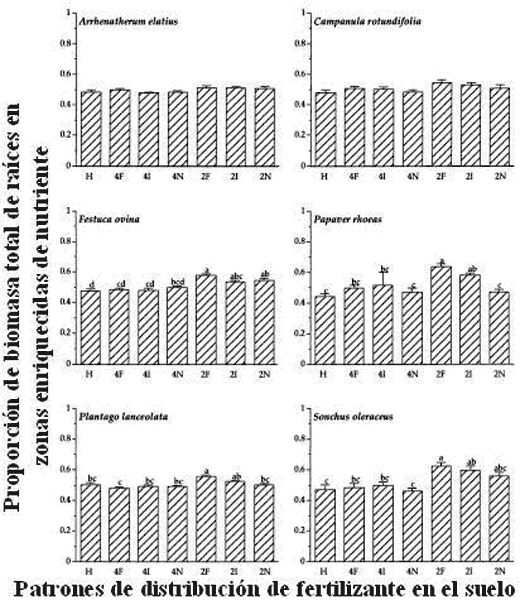
Figura 13. Patrones de distribución de raíces de seis especies (Arrhenatherum elatius, Festuca ovina, Plantago lanceolata, Campanula rotundifolia, Papaver rhoeas y Sonchus oleraceus) en tratamientos de Fertilizante: H (Homogéneo) ó diferentes ubicaciones en la maceta (Dushyantha y col., 2001)
Modificaciones en el patrón de crecimiento de las raíces y en la absorción de fosfato puede en parte compensar el suministro deficiente de fosfato del resto del sistema radicular. Luego de un período de retraso en el crecimiento por las condiciones de estrés de fósforo, la concentración del mismo en la planta y la velocidad de crecimiento relativo se restablecen a niveles que igualan a las plantas testigos (Drew y Saker, 1978).
No todas las especies pueden considerarse con la capacidad de incrementar su sistema radicular en zonas del suelo ricas en nutrientes. Trabajos de Campbell y col. (1991) y Dushyantha y col. (2001), muestran que aquellas especies que naturalmente poseen un sistema radicular muy desarrollado, poseen una menor capacidad de desarrollar selectivamente raíces en zonas del suelo en los que el nivel de fósforo presenta mayor concentración del nutriente (Fig. 13). Este hecho sugiere mecanismos fisiológicos que diferencian a las especies en su capacidad de exploración y está de acuerdo con Collar y col. (2006) y los trabajos de Neumann y Römheld (1999) (Fig. 14).
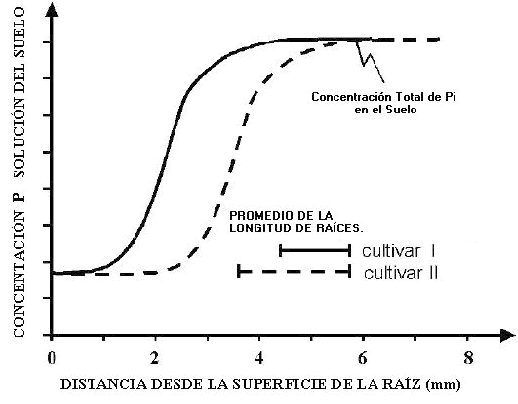
Figura 14. Disminución del nivel de Pi en la rizósfera en relación a las diferencias genotípicas en longitud de los pelos radicales (Neumann y Römheld, 1999)
Anghinoni y Barber (1980) demuestran que los cambios en la masa radicular puede afectar positivamente la superficie de absorción radicular (Tabla 1). Se observa que en la medida que se incrementa el déficit de Pi, se incrementa la biomasa y la longitud radicular, en tanto que disminuye el diámetro de las raíces, indicación de producción de raíces más finas y como consecuencia un aumento de la superficie activa de la raíz. En tanto que una disminución de la relación Parte aérea/Raíz puede implicar un menor requerimiento de nutriente por la planta.
Tabla 1. Crecimiento de parte aérea y raíces de plantas de maíz 12 días pos germinación, en respuesta al aumento de la duración de la deficiencia de P (Anghinoni y Barber, 1980)
| Días sin Suministro de P | Biomasa Total Parte Aérea (g MS Maceta-1) | Raíz Biomasa Total (g MS Maceta-1) | Longitud Total Raíces (m Maceta-1) | Diámetro Raíces (mm) |
| 1 | 2.10 | 0.27 | 46.4 | 2.27 |
| 2 | 2.34 | 0.31 | 57.7 | 2.23 |
| 4 | 1.93 | 0.40 | 75.7 | 1.99 |
| 6 | 1.65 | 0.43 | 90.8 | 1.84 |
Los resultados muestran que los cambios en la morfología modifican la actividad del sistema radical y produce cambios en la capacidad de competencia intra o interespecifica de las plantas, contribuyendo al mantenimiento de la diversidad de las especies.
MODIFICACIONES BIOQUÍMICAS EN LA RIZOSFERA
En muchos suelos, la absorción del fósforo puede estar severamente limitada por la casi nula solubilidad de compuestos en forma orgánica del P tales como, Ca / Mg y Fe / Al fitatos (Adams y Pate, 1992).
Exudados de las raíces en la rizósfera provocan cambios en la zona de absorción.
Estos exudados puede agruparse en tres tipos (Marschner , 1986):
- Sustancias de bajo peso molecular, como ácidos orgánicos, azúcares, aminoácidos y compuestos fenólicos.
- Mucílagos de alto peso molecular, principalmente polisacáridos y ácidos poligalacturonicos.
- Células ó restos de tejidos muertos y productos de su degradación.
En la medida que la competencia entre raíces se incrementa, se genera un ecosistema de baja disponibilidad de Pi, debido a que decrece la difusión como consecuencia de una disminución del gradiente de concentración iónica entre la rizósfera y el suelo. En estas condiciones, muchas plantas y especialmente las adaptadas a condiciones de bajos niveles de fosfato han desarrollado mecanismos bioquímicos para solubilizar compuestos complejos de Pi. Estas especies poseen la capacidad de liberar en la rizosfera ácidos orgánicos (malato, citrato y oxalato), azúcares, aminoácidos, fitosideróforos, polifenoles, vitaminas y hormonas. Así, puede observarse que las leguminosas son muy eficientes en la utilización de roca fosfatada como fuente de Pi, dado justamente por la liberación de ácidos orgánicos (Grayston y col., 1996; Neumann y Römheld, 2000; Hinsinger , 2001; Shane y Lambers, 2005).
El trabajo de Numann y col. (1999) con plantas de trigo (Triticum aestivum L. cv Haro), tomate (Lycopersicon esculentum L. cv Moneymaker), garbanzo (Cicer arietinum) y lupino ( Lupinus albus L. cv Amiga) (Fig. 15) sometidas a deficiencias de fósforo, muestra que en condiciones de estrés nutricional, las especies tienen diferentes respuestas en relación a la liberación de compuestos orgánicos , los cuales están asociados a la capacidad de absorber Pi en condiciones de baja disponibilidad del mismo en el suelo. El ácido cítrico fue uno de los principales ácidos orgánicos acumulados en las raíces de todas las especies investigadas, en respuesta a la deficiencia de Pi y esto se asoció con aumento de la actividad de la enzima PEP-carboxilasa, que se requiere para la biosíntesis de citrato. En estos casos resultó un factor determinante de la absorción del fósforo y en el crecimiento de las plantas.
Los resultados sugieren que la deficiencia de P induce la exudación de ácidos carboxílicos desde los tejidos de la raíz y depende de la capacidad de acumulación de los mencionados tejidos. La capacidad de acumular ácidos carboxílicos está regulada por la velocidad de biosíntesis, velocidad de degradación y la compartimentalización de los mismos ó la de sus precursores.
La exudación de gran cantidad de ácido en el caso del Lupino blanco, se ha relacionado con mecanismos específicos de transporte mediado por canales iónicos. En todos los casos de regulación el incremento de exudación de ácido cítrico es mediado por una señal de deficiencia interna de Pi en la raíz (Fig. 15).
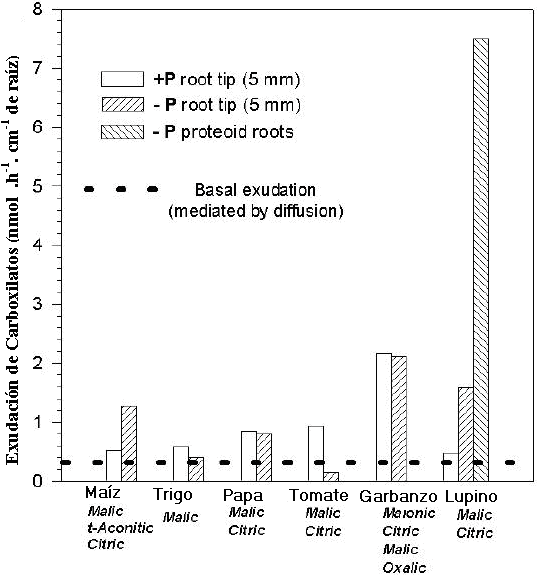
Figura 15. Efecto de la deficiencia de P en la exudación de carboxilatos en las zonas apical de la raíz en plantas de diferentes especies (Neumann y Römheld. 1999).
Dadas las evidencias de la exudación de citrato por la raíz y concomitantemente la mayor absorción de Pi por la misma, Kirk (1999) (Fig. 15) propone que el principal mecanismo de solubilización de P del suelo por el citrato se debe a:
1. la quelación de los metales involucrados en la fijación de los iones P, lo cual está de acuerdo con los trabajos posteriores realizados por Gerke y col. (2000a)
2. la formación de un complejo soluble en citrato metal-P como ya había sido propuesto por Gardner y col. (1983) en sus trabajos con lupino blanco.
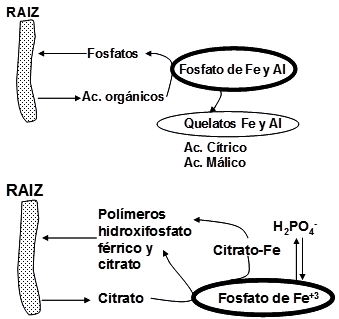
Figura 16. Representación esquemática de los mecanismos en la rizósfera que conllevan a la disponibilidad de P mediante la exudación de diferentes compuestos por la raíz
El fósforo orgánico insoluble y por tanto no disponible para las plantas, puede encontrarse entre el 20 - 80% del total del suelo (Tarafdar y Marschner, 1994). A nivel de células y tejidos de las raíces de las plantas, en condiciones de deficiencia de Pi, los mecanismos por los cuales se produce una hipersecreción de compuestos (fosfohidrolasas, fosfatasas ácidas, nucleasas, fitasas, hidrogeniones, ácidos orgánicos y otros) se presenta en forma de esquema (Fig. 16). Los mismos facilitan la disponibilidad de Pi en la rizosfera de la planta. La capacidad de secreción inducida a nivel de la raíz, varía de manera considerable entre las especies e incluso entre los cultivares (Li y col., 1997b; Römer y col., 1995). La secreción de fosfatasas ácidas es probablemente regulada a nivel transcripcional e incluye mecanismos de detección de concentraciones de fósforo externo en el medio de cultivo y la inducción diferencial de isoenzimas (Wasaki y col., 1999). La deficiencia externa de Pi desencadena un mecanismo a través de canales anionicos de transporte, de liberación citrato hacía el exterior, lo cual está acoplado a una bomba de H +-ATPasa. Este mecanismo es auto regulado por procesos de retroalimentación que evita las concentraciones de citrato a niveles no fisiológicos.
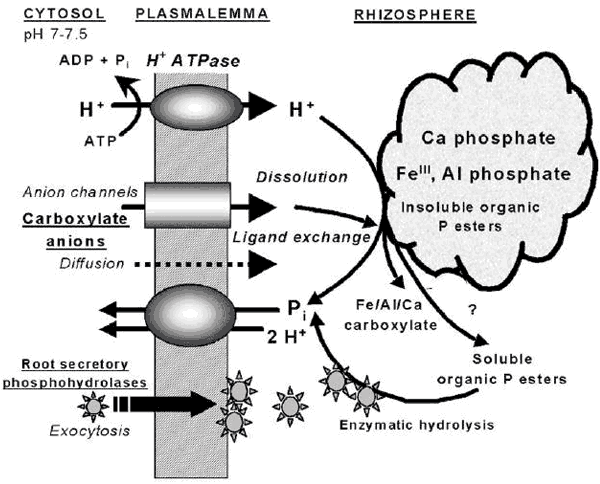
Figura 17. Modelo de los mecanismos internos de la raíz, de liberación H+, ácidos carboxílicos y fosfohidrolasas inducida por señales de bajo nivel de Pi del suelo y como consecuencia la movilización y disponibilidad de fósforo desde fuentes de baja disponibilidad.
Liu y col. (2005), demostraron la interrelación de este mecanismo bioquímico con procesos como la fotosíntesis, dado que una interrupción del suministro de azúcares vía floema a las raíces, se relaciona con la inducción de señales de deficiencia de P en Lupinus albus (Fig. 14). La complejidad del sistema en la expresión de plantas P-deficiente, involucra no solo los mencionados compuestos de exudación, sino que además se producen cambios en los niveles de fitohormonas, fundamentalmente auxinas, citoquininas, etileno y ABA (Fig. 18) (Hammond y White, 2008).
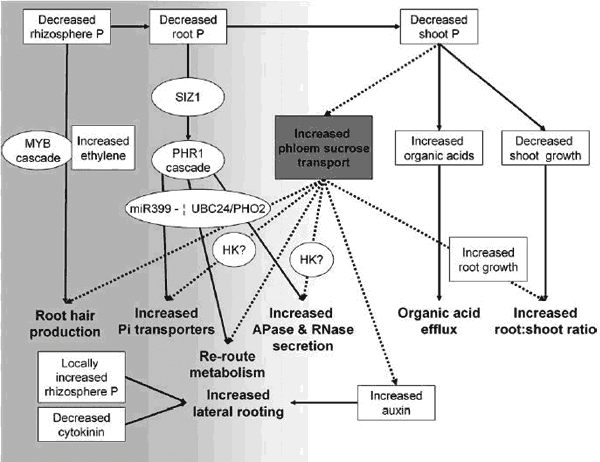
Figura 18. Complejidad del sistema de cascadas de señalización que tienen como respuesta cambios en la morfología del sistema radical, cambios en la capacidad de transporte Pi, la exudación de ácidos orgánicos y enzimas en la rizosfera (Hammond y White, 2008)
Si bien los mecanismos no son universales y varían con las especies y variedades, una simplificación de las respuestas de las plantas a la deficiencia de fósforo se presenta en la Fig. 19. Puede observarse que para aumentar la disponibilidad y el suministro de Pi ante una señal de deficiencia de P, las plantas deben adaptar la bioquímica, la fisiología y la morfología de sus sistemas radiculares, como forma de maximizar la absorción de Pi (Vance y col., 2003). El incremento en la proliferación de los pelos radiculares tiene como consecuencias un aumento de la superficie de absorción que puede superar el 70% a las plantas testigo, la cual es responsable del mayor volumen de suelo explorado (Bates y Lynch, 1996). Otra respuesta a la deficiencia de Pi, consiste en la alteración de la morfología del sistema radicular, lo cual permite incrementar la capacidad de reservas y equilibrio de la planta, incrementando la relación raíz:parte aérea (Lynch, 1995). Los ácidos carboxílicos y enzimas exudados por la raíz, permiten la liberación de fósforo desde fuentes orgánicas e inorgánicas (Neumann y col., 2000). Las raíces de ciertas plantas poseen la capacidad de establecer asociaciones microrríticas como una alternativa para el incremento de la superficie de absorción de fósforo y la solubilización mediante fosfatasas, de formas no disponibles de fósforo. Así especies como Lupino no establecen asociaciones con micorrizas en tanto otras como Cucurbita pepo (Cucurbitaceas) pueden beneficiarse de este mecanismo asociativo (Lambers y col., 2006).
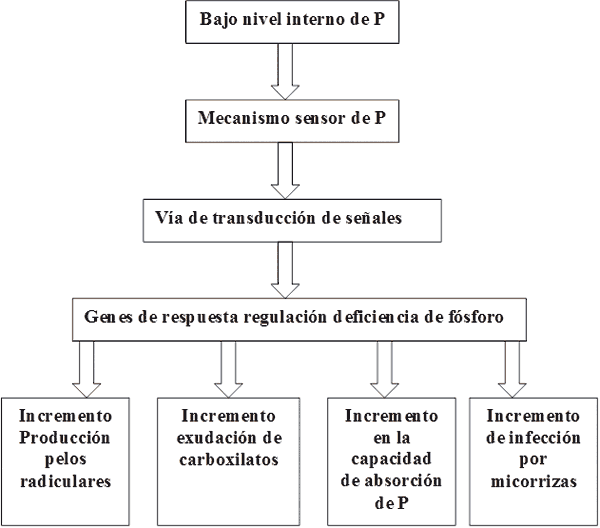
Figura 19. Respuestas de las plantas a la deficiencia de fósforo inorgánico (Pi). Una baja disponibilidad externa de Pi disminuye nivel de P interno en la planta, se desencadenan una serie de respuestas mediada por diferentes señales, que depende de la especie y la variedad considerada (Lambers et al., 2006).
CONCLUSIONES
La deficiencia de fosfato en muchos ecosistemas agrícolas, conlleva que se establezca una competencia por el nutriente Pi. Es este elemento uno de los principales factores que limitan el crecimiento de la planta, por lo que los mecanismos de adaptación de especies y variedades, juega un rol fundamental en la supervivencia y la productividad del cultivo. Entender la fisiología de la raíz, los mecanismos involucrados en la absorción y competencia por el fósforo es fundamental cuando se piensa en pasturas consociadas.
A pesar de los trabajos pioneros de Epstein (1966) u otros investigadores (Drew y Nye, 1963), el paradigma de la “revolución verde” en la década de los 60, trató de adaptar el suelo a la planta. En esos años las curvas de respuesta de producción de biomasa, en función de la dosis de fertilizante aplicado, ocuparon gran parte de las horas de investigación y páginas de publicaciones periódicas. Ante una gran disponibilidad de fósforo para la planta, los mecanismos por los cuales una planta puede competir con otras por el recurso fósforo, quedaron enmascarados.
Los posibles factores y procesos morfológicos, fisiológicos y bioquímicos, mediante los cuales las raíces pueden alterar la química y la movilidad del nutriente y por tanto la biodisponibilidad de fósforo inorgánico del suelo en el rizósfera, cubre un amplio espectro de situaciones tanto a nivel de especies y genotipos como de situaciones del hábitat. Es así que las aproximaciones posteriores a la “revolución verde” se focalizan fundamentalmente en los mecanismos de la absorción del fósforo y el incremento de la eficiencia en la utilización del nutriente por las planta (Lynch, 1998).
Considerando los diferentes aspectos tratados, por los cuales las raíces pueden competir en el hábitat, un simple enfoque de: nivel de fósforo aplicado al suelo vs. curva de respuesta, queda en un segundo plano de interés. Por otra parte los avances de la fisiología vegetal, la bioquímica y la biología molecular contribuyendo al área de la ecofisiología, permiten enfocar la aproximación de competencia por el fósforo desde el punto de vista de “adaptar la planta al suelo”.
Raíces con la capacidad de incrementar la superficie de absorción, crecimiento localizado en zonas de suelo, adaptación ó cambios de los parámetros cinéticos de la absorción, la secreción de compuestos por la rizósfera que logren una mayor disponibilidad del Pi, son algunos de los cambios que pueden conferir ventajas competitiva de las raíces en un hábitat de bajo contenido de nutriente.
Es necesario dejar atrás el enfoque reduccionista de la “revolución verde”, comprender que se debe avanzar a los enfoques multidisciplinarlos con la botánica, bioquímica, fisiología vegetal, biología molecular, edafología y la ecofisiología como vía fundamental de lograr avances en “adaptar la planta al suelo”.